Theories Behind Plant Tissue Culture
Basics of Plant Tissue Culture
Yang Yen
College of Agriculture and Biological Sciences at South Dakota State University, USA
 Lesson Navigation Tips: Lesson Navigation Tips:
- Click on ’Animations’ button found to the left in order to view the animation which supplements this lesson. You can also click on the animation icon within the text.
- Click once on figures to see enlarged versions.
- Click once on words in color to bring up their definitions.
|
After delivering a foreign gene into a target genome, you need to bring the transgenic cell to a complete plant. This step has to be done by plant tissue culture. Plant tissue culture is a biotechnique based on the promise that an organ, tissue or cell of a plant can be in vitro manipulated to grow back in to a complete plant. Therefore, plant tissue culture is the foundation and in most cases the bottle-neck step for plant genetic engineering.
The history of plant tissue culture can be traced back to near the turn of the 20th century when Gottlieb Haberland reported his culture of leaf mesophyll tissue and hair cells, though the cultured cells did not divide ( see Steward 1968). Haberland’s student Kotte (1922) reported in vitro growth of isolated root tips. White (1934) repeatedly reported subculture of root tip-derived tissues of tomato. Techniques for plant tissue culture progressed rapidly during the 1930s due to the discovery of the necessity of B vitamins and auxin for the growth of isolated meristem tissues. The works of Skoog and his associates on the nutritional requirements of tobacco tissue culture led to not only the discovery of plant growth hormones, kinetin and cytokinins, but also to the formation of an important plant tissue culture medium, the MS medium (Skoog and Tsui, 1951; Skoog and Miller, 1957; Murashige and Skoog, 1962). Since the 1960s, tissue and cell culture has increasingly been used as a tool by plant scientists and biotechnologists. Yet, much remains to be explored in terms of methodology, procedures and the theories behind.
In this lecture, we will learn the basic theories which guide our practice, and the techniques that are fundamental for plant tissue culture. The two parts can be learned separately.
Development of this lesson was supported in part by Cooperative State Research, Education, & Extension Service, U.S. Dept of Agriculture under Agreement Number 98-EATP-1-0403 administered by Cornell University and the American Distance Education Consortium (ADEC).
Any opinions, findings, conclusions or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the view of the U.S. Department of Agriculture.Cell Totipotency
A plant grows by increasing its cell population while the cells specialize their functions. Increasing cell population is done by cell division (also called mitosis). Before a mother cell divides into two daughter cells, it makes an exact copy of its genome first. As a result, the two daughter cells usually have exactly the same genetic makeup as their mother cell. Therefore, every living cell of a plant should contain all the genes the plant has and thus has the capacity to grow back to a full plant. This is called cell totipotency .
The process of specializing cells’ functions is called cell differentiation . It is accompanied by morphogenesis, the change of the cells’ morphology. Differentiation is done by turning on certain genes and turning off some others at a certain time. Therefore, for a highly differentiated cell to grow into a full plant, the differentiation process has to be reversed (called de-differentiation ) and repeated again ( called re-differentiation). Theoretically, all living cells can revert to an undifferential status through this process. However, the more differentiated a cell has been, the more difficult it will be to induce its de-differentiation. Practically, the younger or the less differentiated a cell is, the easier to culture it into a full plant. The ease of fulfilling the cell totipotency also varies tissue by tissue, genotype by genotype and species by species. Genotype dependency is often the bottle-neck in plant tissue culture and also in plant genetic engineering.
Organogenesis
Click here to see an Animation of Organogenesis An adult plant consists of many specialized cell organizations: tissues and organs. Tissues, such as meristem, cortex, phloem and epidermis, consist of cells of uniform shape and specialized function. Several tissues are organized together to form an organ, such as leaves, roots, flowers and the vascular system. The process of initiation and development of an organ is called organogenesis. In plant tissue culture, inducing organogenesis is an important way to regenerate plants from the culture.
Organogenesis in plant tissue culture involves two distinct phases: dedifferentiation and redifferentiation. Dedifferentiation begins shortly after the isolation of the explant tissues with an acceleration of cell division and a consequent formation of a mass of undifferentiated cells (called callus). Redifferentiation, also called budding in plant tissue culture, may begin any time after the first callus cell forms. In this process of tissue called organ primordia is differentiated from a single or a group of callus cells. The organ primordia give rise to small meristems with cells densely filled with protoplasm and strikingly large nuclei. The development (or growth) of an organ is monopolar . Polarity of the longitudinal axis of the organizing growing points of the organs can be seen some time after the formation of meristem tissues. Different types of specialized cells again differentiate. The vascular system is formed connecting the new organs to their parent explants or callus mass.
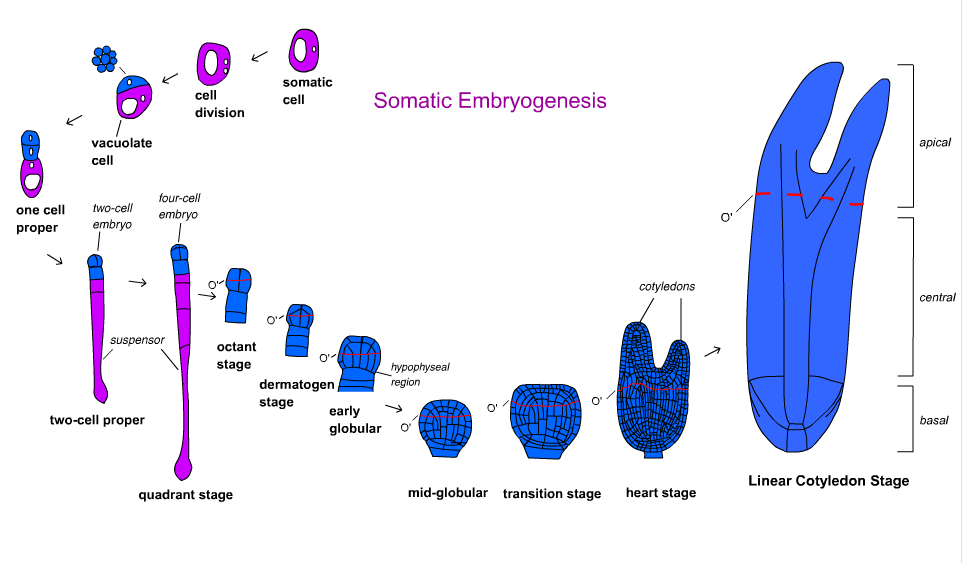
Somatic Embryogenesis
Click here to see an Animation of Somatic Embryogenesis The process of formation of an embryo is called embryogenesis. Embryogenesis starts from a single embryogenic cell, that can be a zygote (the product of the fusion of an egg and a sperm during fertilization), or an undifferentiated callus cell. Embryos developing from zygotes are called zygotic embryos, while those derived from somatic cells are called somatic embryos. During the embryonic development, the polar axis of the plant is established, domains that set up the organization of the plant body are defined, and the primary tissue and organ systems are delineated. Somatic embryogenesis is another important way to regenerate new plants in plant tissue culture.
Embryo development occurs through an exceptionally organized sequence of cell division, enlargement and differentiation. Zygotic and somatic embryos share the same gross pattern of development. Both types of embryos develop as passing through typical developmental stages . Embryo development is bipolar, having a shoot pole and a radicular pole at opposite ends. Embryos are not organs because they are structurally independent from their parent body (i.e. they do not have a vascular system connecting them with their parent plant body).

| Progenitor cell undergoes unequal division. |
The physical, observable transition from a nonembryogenic cell to an embryogenic cell in somatic embryogenesis appears to occur when the progenitor cell undergoes an unequal division, resulting in a larger vacuolate cell and a small, densely cytoplasmic (embryogenic) cell . The embryogenic cell then either continues to divide irregularly to form a proembryonal complex or divides in a highly organized manner to form a somatic embryo.

| Somatic Embryo Chart |
Somatic embryos usually do not mature properly. Instead, due to environmental factors such as constant contact with inducing medium, somatic embryos often deviate from the normal developmental pattern by bypassing embryo maturation producing callus, undergoing direct secondary embryogenesis and/or germinating precociously. Somatic embryos growing from proembryonal complexes tend to develop asynchronously so that several stages are present in culture at any given time.
Culture Media
Culture medium is the most important part of plant tissue culture. A successful plant tissue culture system largely relies on a right culture medium formula. Plant tissue culture media usually contains inorganic elements, organic compounds and a support matrix. Culture media provides the cultures the necessary inorganic nutrients that are usually available from soil. In addition, they also provide the cultures the necessary organic compounds such as vitamins, and carbon source, which are usually produced in plants. Sometimes, plant growth regulators are added to the medium to stimulate cell division and/or differentiation.
Another important function of a culture medium is creating a necessary environment for plants to grow. For example, solid media, functioning like the soil, provides a physical support for the cultures to keep contact with air for respiration, and for regenerated plantlets to root into. A liquid medium enables explants to keep constant, maximum contact with nutrient supplies. A selective reagent may also be included in a culture medium to restrict the growth of certain cultures. Therefore, medium formulas vary depending upon the purpose of plant tissue culture. Different media may also be used during a plant tissue culture process.
Mineral Elements, Macro
Based on the amount needed, mineral elements are usually classified into two groups: macroelements and microelements. Macroelements consist of elements in large supply, while microelements are usually only needed in trace amount.
The Macroelements
Calcium (Ca): Calcium functions as a cofactor with many enzymes and is particularly important in cell well synthesis. Calcium deficiency may result in shoot tip necrosis. Calcium used in plant tissue culture is mostly in the forms of calcium chloride and calcium nitrate in a concentration of 1-3 mM.
Magnesium (Mg): Magnesium is critical for enzyme functioning, and an integral component of chlorophyl molecule. Magnesium is also a cation in plant to balance negative ions. The most used form of magnesium in plant tissue culture is magnesium sulfate in a concentration of 1-3 mM.
Nitrogen (N): Nitrogen is essential to plant life. It is a constituent of many important molecules, including amino acids, proteins, nucleic acids, certain hormones and chlorophyll. The nitrate ion (NO3- oxidized) and the ammonium ion (NH4+ reduced) are the two nitrogen forms that are mostly used in plant tissue culture. Nitrate is usually added at 25-40 mM and ammonium at 2-20 mM with a total amount of inorganic nitrogen at 25-60 mM. Many plants appear to grow best if given both forms. Nitrogen may also be given in organic forms as amino acids, hydrolysates and organic acids. Organic forms of nitrogen cannot totally replace inorganic forms.
Phosphorus (P): Phosphorus is an integral part of nucleic acids and other structural compounds in plants. Phosphorus is supplied in culture media as phosphate (PO4-) in a form of sodium or potassium hydrogen phosphate at 1-3 mM.
Potassium (K): Potassium is major positive ion in plants to balance negative ions. The amount of potassium needed in a plant tissue culture medium varies depending on the plant species. Usually it is supplied at 20-30 mM in correlation with that of nitrate.
Sulfur (S): Sulfur is important in protein structure where disulphide bonds between neighboring cystein and methionine residues contribute to the tertiary structure. Sulfur is also a constituent of the vitamins thiamine and biotin and of coenzyme A, an important component in respiration and fatty acid metabolism. In the form of iron-sulfur proteins it is important in electron transfer reactions of photosynthesis. Sulfur is supplied as the SO4- ion usually with magnesium as the cation at 1-3 mM.
Iron (Fe): Iron is required for chlorophyll synthesis as well as in many oxidation/ reduction reactions. Iron usually present in media at 1 ìM. Since iron forms insoluble compounds in alkaline pH, being unavailable to plant in culture, medium pH is critical for iron supply. Fe-EDTA is often used in culture media and enable iron to be available to cultures over a wider pH range than other agents.
Mineral Elements, Micro
The Microelements
The micro elements included in culture media and their metabolic role include the following.
Boron (B): Boron is essential in enzyme activity in lignin biosynthesis and metabolism of phenolic acids. Boron deficiency inhibits cell division and elongation in both root tip and shoot tip meristems. In culture media, boron is supplied as boric acid.
Cobalt (Co): Cobalt is not considered as an essential element by plant physiologists. However, about 0.1 ìM cobalt is widely used in plant culture media. It may be toxic at higher concentrations.
Copper (Cu): As a plant nutrient, copper seems to function primarily as a cofactor for a variety of oxidative enzymes. Copper is used in the form of copper sulfate at 0.1 ìM in culture media. Higher concentration may be toxic.
Iodine (I): The effect of iodine varies depending on plant species. I is not an essential element but added to media to improve growth of roots and callus in culture.
Manganese (Mn): As a cofactor, manganese is required by some enzyme reactions, particularly in respiration and photosynthesis processes. Usually, 5-30 ìM manganese is added to the medium in the form of manganese sulfate.
Molybdenum (Mo): Molybdenum is a cofactor with two enzymes involved in transforming nitrate to ammonium. It is supplied as sodium molybdate at 1 ìM in media.
Zinc (Zn): Zinc is absorbed as the divalent cation Zn2+ by plants. It is an activator of a large number of enzymes. Typical symptoms of zinc deficiency include shortened internodes and smaller leaves. Zinc sulfate is the most common form added to culture media at 5-30 ìM.
Organic Compounds
Organic compounds are added in large quantities to culture media.
Sugars: Sugars serve as an energy source for plant culture. Sucrose is the most commonly used sugar in plant culture media, although glucose, fructose and sorbitol are also used in some medium formulas. Sucrose is partially broken down into fructose and glucose during autoclaving . Sucrose also acts as an osmoticum in the medium. Generally 20-40 gram sucrose is added in each liter of liquid medium. Starches are also used in some medium formulas both as the support substance and energy source.
Vitamins: Vitamins can improve, but are not absolutely necessary for plant growth in culture. Off all vitamins, only thiamin (B1) is considered to be essential for many plant species in culture. It is required for carbohydrate metabolism and the biosynthesis of some amino acids. Nicotinic acid (niacin) and pyridoxine (B6), biotin, folic acid, ascorbic acid (vitamin C) and tocopherol (vitamin E) are also used in some media.
Myo-inositol: Myo-inositol is a sugar alcohol and is thought to be important in membrane and cell wall development. It is usually added to media for monocots as well as some dicots and gymnosperms.
Complex organic Compounds: Complex organic compounds such as coconut milk or juice, yeast extract, fruit juices and fruit pulps are used in some medium formulas. It is not known exactly what components in these organic supplements is responsible for the improved growth of the culture.
Support Substances
Supporting matrix is often required to keep explants from being submerged in the medium. A support matrix may be formed by solidification of a gelling agent, such as agar, agarose and gelatin, or by mechanical support materials, such as filter paper or a polypropylene membrane raft. The selection of a gelling agent is often empirical. A major concern is the degree of hyperhydricity induced. Also for some unknown reasons, a species may grow more vigorously on one gelling agent than on others.
Agar: Agar is the most commonly used gelling agent in plant tissue culture. It is a mixture of polysaccharides derived from red algae. Agar melts at about 100oC and solidifies at about 45oC. Agar does not react with medium components nor is it digested by plant enzymes. Agar does not gel well under acidic conditions (pH < 4.5). All agar contains impurities, the amount and purity of which differ among manufacturers. However, these impurities usually do not interfere with the culture.
Agarose: Agarose is extracted from agar. It does not contains agaropectin and its sulfate groups. Agarose has higher gel strength than agar. Agarose is often used when the impurities in agar are not desired, such as in protoplast and anther culture.
Gellan Gums: Gellan gums such as Gelrite and Phytagel consist of a polysacchraride produced by bacterium Pseudomonas elodea. They are clear and thus makes detection of contamination easier than using agar. Gellan gums will not gel if concentration of divalent cations such as calcium and magnesium is lower than 4 mM or higher than 8 mM.
Plant Growth Regulators
Plant growth regulators (also called plant hormones) are numerous chemical substances that profoundly influence the growth and differentiation of plant cells, tissues and organs. Plant growth regulators function as chemical messengers for intercellular communication . There are currently five recognized groups of plant hormones: auxins, gibberellins, cytokinins, abscisic acid (ABA) and ethylene. They work together coordinating the growth and development of cells. Ethylene is mainly involved in abscission and flower secscence in plants and is rarely used in plant tissue culture. In addition to the five principal growth regulators, two other groups sometimes appear to be active in regulating plant growth, the brassinosteroids and polyamines.
Auxins: Auxins stimulate cell elongation and influence a host of other developmental responses, such as root initiation, vascular differentiation, tropic responses, apical dominance and the development of auxiliary buds, flowers and fruits. Auxins are synthesized in the stem and root apices and transported through the plant axis. The principal auxin in plants is indole-3-acetic acid (IAA). Several other indole derivatives, all as precursors to IAA, are known to express auxin activity, probably by converting to IAA in the tissue. Auxins in plant tissue culture are used to induce callus from explants , and cause root and shoot morphogenesis . Auxins are often most effective in eliciting their effects when combined with cytokinins.
Cytokinins: Cytokinins are able to stimulate cell division and induce shoot bud formation in tissue culture. They usually act as antagonists to auxins. (Cytokinins are N6 substituted derivatives of the nitrogenous purine base adenine.) Cytokinins most used in tissue culture include zeatin, adenine, 6-(g,g-dimethylallylamino)purine (2 iP) and kinetin. Cytoknins often inhibit embryogenesis and root induction.
Gibberellins: The main effect of gibberellins in plants is to cause stem elongation and flowering. They are also prominently involved in mobilization of endosperm reserves during early embryo growth and seed germination. Gibberellins are an extensive chemical family based on the ent-gibberellane structure. There exit over 80 different gibberellin compounds in plants but only giberrellic acid (GA3) and GA4+7 are often used in plant tissue culture. In tissue culture, gibberellins are used to induce organogenesis, particularly adventitious root formation.
Abscisic Acid: Abscisic acid (ABA) in plants is a terpenoid involved primarily in regulating seed germination, inducing storage protein synthesis and modulating water stress. In plant tissue culture, it is used to help somatic embryogenesis, particularly during maturation and germination.
Ethylene: Ethylene is a simple gaseous hydrocarbon with the chemical structure H2C=CH2. Ethylene is apparently not required for normal vegetative growth. However, it can have a significant impact on development of root and shoots. Usually, ethylene is not used in plant tissue culture.